рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Медицина
- /
- Механизм влияния гормонов на организм
Реферат Курсовая Конспект
Механизм влияния гормонов на организм
Механизм влияния гормонов на организм - раздел Медицина, Гены и ДНК Существуют Два Основных Механизма Действия Гормонов На Уровне Клетки: ...
Существуют два основных механизма действия гормонов на уровне клетки:
1. Реализация эффекта с наружной поверхности клеточной мембраны.
2. Реализация эффекта после проникновения гормона внутрь клетки.
1)Реализация эффекта с наружной поверхности клеточной мембраны
В этом случае рецепторы расположены на мембране клетки. В результате взаимодействия гормона с рецептором активируется мембранный фермент — аденилатциклаза. Этот фермент способствует образованию из аденозинтрифосфорнои кислоты (АТФ) важнейшего внутриклеточного посредника реализации гормональных эффектов — циклического 3,5-аденозинмонофосфата (цАМФ). цАМФ активирует клеточный фермент протеинкиназу, реализующую действие гормона. Установлено, что гормоно-зависимая аденилатциклаза — это общий фермент, на который действуют различные гормоны, в то время как рецепторы гормонов множественны и специфичны для каждого гормона. Вторичными посредниками кроме цАМФ могут быть циклический 3,5-гуанозинмонофосфат (цГМФ), ионы кальция, инозитол-трифосфат. Так действуют пептидные, белковые гормоны, производные тирозина — катехоламины. Характерной особенностью действия этих гормонов является относительная быстрота возникновения ответной реакции, что обусловлено активацией предшествующих уже синтезированных ферментов и других белков.
Гормоны осуществляют свое биологическое действие, комплексируясь с рецепторами – информационными молекулами, трансформирующими гормональный сигнал в гормональное действие. Большинство гормонов взаимодействуют с рецепторами, расположенными на плазматических мембранах клеток, а другие гормоны – с рецепторами, локализованными внутриклеточно, т.е. с цитоплазматическими и ядерными.
Плазматические рецепторы в зависимости от структуры подразделяются на:
1. рецепторы, трансмембранный сегмент которых состоит из семи фрагментов (петель);
2. рецепторы, трансмембранный сегмент которых состоит из одного фрагмента (петли или цепи);
3. рецепторы, трансмембранный сегмент которых состоит из четырех фрагментов (петель).
К гормонам, рецептор которых состоит из семи трансмембранных фрагментов, относятся:
АКТГ, ТТГ, ФСГ, ЛГ, хорионический гонадотропин, простагландины, гастрин, холецистокинин, нейропептид Y, нейромедин К, вазопрессин, адреналин (a-1 и 2, b-1 и 2), ацетилхолин (М1, М2, М3 и М4), серотонин (1А, 1В, 1С, 2), дофамин (Д1 и Д2), ангиотензин, вещество К, вещество Р, или нейрокинин 1, 2 и 3 типа, тромбин, интерлейкин-8, глюкагон, кальцитонин, секретин, соматолиберин, ВИП, гипофизарный аденилатциклазактивирующий пептид, глютамат (MG1 – MG7), аденин.
Ко второй группе относятся гормоны, имеющие один трансмембранный фрагмент:
СТГ, пролактин, инсулин, соматомаммотропин, или плацентарный лактоген, ИФР-1, нервные факторы роста, или нейротрофины, фактор роста гепатоцитов, предсердный натрийуретический пептид типа А, В и С, онкостатин, эритропоэтин, цилиарный нейротрофический фактор, лейкемический ингибиторный фактор, фактор некроза опухолей (р75 и р55), нервный фактор роста, интерфероны (a, b и g), эпидермальный фактор роста, нейродифференцирующий фактор, факторы роста фибробластов, факторы роста тромбоцитов А и В, макрофагный колониестимулирующий фактор, активин, ингибин, интерлейкины-2, 3, 4, 5, 6 и 7, гранулоцито-макрофагный колониестимулирующий фактор, гранулоцитный колониестимулирующий фактор, липопротеин низкой плотности, трансферрин, ИФР-2, урокиназный плазминогенный активатор.
К гормонам третьей группы, рецептор которых имеет четыре трансмембранных фрагмента, относятся:
ацетилхолин (никотиновые мышечные и нервные), серотонин, глицин, g-аминомасляная кислота.
Cопряжение рецептора с эффекторными системами осуществляется через так называемый G-белок, функция которого заключается в обеспечении многократного проведения гормонального сигнала на уровне плазматической мембраны. G-белок в активированной форме стимулирует через аденилатцик-лазу синтез циклического АМФ, который запускает каскадный механизм активирования внутриклеточных белков.
Общим фундаментальным механизмом, посредством которого реализуются биологические эффекты «вторичных» мессенджеров внутри клетки, является процесс фосфорилирования –дефосфорилирования белков при участии широкого разнообразия протеинкиназ, катализирующих транспорт концевой группы от АТФ на ОН-группы серина и треонина, а в ряде случаев – тирозина белков-мишеней. Процесс фосфорилирования представляет собой важнейшую посттрансляционную химическую модификацию белковых молекул, коренным образом изменяющую как их структуру, так и функции. В частности, он вызывает изменение структурных свойств (ассоциацию или диссоциацию составляющих субъединиц), активирование или ингибирование их каталитических свойств, в конечном итоге определяя скорость химических реакций и в целом функциональную активность клеток.
Аденилатциклазная мессенджерная система
Наиболее изученным является аденилатциклазный путь передачи гормонального сигнала. В нем задействовано мимимум пять хорошо изученных белков:
1)рецептор гормона;
2)фермент аденилатциклаза, выполняющая функцию синтеза циклического АМФ (цАМФ);
3)G-белок, осуществляющий связь между аденилатциклазой и рецептором;
4)цАМФ-зависимая протеинкиназа, катализирующая фосфорилирование внутриклеточных ферментов или белков-мишеней, соответственно изменяя их активность;
5)фосфодиэстераза, которая вызывает распад цАМФ и тем самым прекращает (обрывает) действие сигнала
Показано, что связывание гормона с β-адренергическим рецептором приводит к структурным изменениям внутриклеточного домена рецептора, что в свою очередь обеспечивает взаимодействие рецептора со вторым белком сигнального пути – ГТФ-связывающим.
ГТФ-связывающий белок – G-белок – представляет собой смесь 2 типов белков:
активного Gs(от англ. stimulatory G)
ингибиторного Gi
В составе каждого из них имеется три разные субъединицы (α-, β- и γ-), т.е. это гетеротримеры. Показано, что β-субъединицы Gs и Gi идентичны; в то же время α-субъединицы, являющиеся продуктами разных генов, оказались ответственными за проявление G-белком активаторной и ингибиторной активности. Гормонрецепторный комплекс сообщает G-белку способность не только легко обменивать эндогенный связанный ГДФ на ГТФ, но и переводить Gs-белок в активированное состояние, при этом активный G-белок диссоциирует в присутствии ионов Mg2+ на β-, γ-субъединицы и комплекс α-субъединицы Gs в ГТФ-форме; этот активный комплекс затем перемещается к молекуле аденилатциклазы и активирует ее. Сам комплекс затем подвергается самоинактивации за счет энергии распада ГТФ и реассоциации β- и γ-субъединиц с образованием первоначальной ГДФ-формы Gs.
Аденилатциклазный путь передачи гормонального сигнала..
Аденилатциклаза представляет собой интегральный белок плазматических мембран, его активный центр ориентирован в сторону цитоплазмы и катализирует реакцию синтеза цАМФ из АТФ:
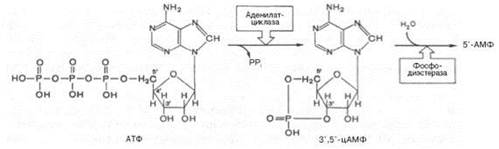
Каталитический компонент аденилатциклазы, выделенный из разных тканей животных, представлен одним полипептидом. В отсутствие G-белков он практически неактивен. Содержит две SH-группы, одна из которых вовлечена в сопряжение с Gs-белком, а вторая необходима для проявления каталитической активности.Под действием фосфоди-эстеразы цАМФ гидролизуется с образованием неактивного 5'-АМФ.
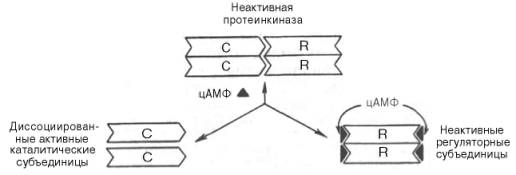
Протеинкиназа – это внутриклеточный фермент, через который цАМФ реализует свой эффект. Протеинкиназа может существовать в 2 формах. В отсутствие цАМФ протеинкиназа представлена в виде тетрамерного комплекса, состоящего из двух каталитических (С2) и двух регуляторных (R2) субъединиц; в этой форме фермент неактивен. В присутствии цАМФ протеинкиназный комплекс обратимо диссоциирует на одну R2-субъединицу и две свободные каталитические субъединицы С; последние обладают ферментативной активностью, катализируя фосфорилирование белков и ферментов, соответственно изменяя клеточную активность.
Гуанилатциклазная мессенджерная система
Довольно долгое время циклический гуанозинмонофосфат (цГМФ) рассматривался как антипод цАМФ. Ему приписывали функции, противоположные цАМФ. К настоящему времени получено много данных, что цГМФ принадлежит самостоятельная роль в регуляции функции клеток. В частности, в почках и кишечнике он контролирует ионный транспорт и обмен воды, в сердечной мышце служит сигналом релаксации и т.д.
Биосинтез цГМФ из ГТФ осуществляется под действием специфической гуанилатциклазы по аналогии с синтезом цАМФ.
Известны четыре разные формы гуанилатциклазы, три из которых являются мембраносвязанными и одна – растворимая открыта в цитозоле.
Получены доказательства, что большинство эффектов цГМФ опосредовано через цГМФ-зависимую протеинкиназу, названную протеинкина-зой G.
Уровень циклических нуклеотидов цАМФ и цГМФ в клетке контролируется соответствующими фосфодиэстеразами, катализирующими их гидролиз до 5'-нуклеотидмонофосфатов и различающимися по сродству к цАМФ и цГМФ. Выделены и охарактеризованы растворимая кальмоду-линзависимая фосфодиэстераза и мембраносвязанная изоформа, не регулируемая Са2+ и кальмодулином.
Са2+-мессенджерная система
Ионам Са2+ принадлежит центральная роль в регуляции многих клеточных функций. Изменение концентрации внутриклеточного свободного Са2+ является сигналом для активации или ингибирования ферментов, которые в свою очередь регулируют метаболизм, сократительную и секреторную активность, адгезию и клеточный рост. Источники Са2+ могут быть внутри- и внеклеточными. В норме концентрация Са2+ в цитозоле не превышает 10-7 М, и основными источниками его являются эндоплазматический ретикулум и митохондрии. Нейрогормональные сигналы приводят к резкому повышению концентрации Са2+ (до 10–6 М), поступающего как извне через плазматическую мембрану (точнее, через потенциалзависимые и рецепторзависимые кальциевые каналы), так и из внутриклеточных источников. Одним из важнейших механизмов проведения гормонального сигнала в кальций–мессенджерной системе является запуск клеточных реакций (ответов) путем активирования специфической Са2+-кальмодулин-зависимой протеинкиназы. Регуляторной субъединицей этого фермента оказался Са2+-связывающий белок кальмодулин. При повышении концентрации Са2+ в клетке в ответ на поступающие сигналы специфическая протеинкиназа катализирует фосфорилирование множества внутриклеточных ферментов – мишеней, регулируя тем самым их активность.
К внутриклеточной системе мессенджеров относят также производные фосфолипидов мембран эукариотических клеток, в частности фосфорилированные производные фосфатидилинозитола. Эти производные освобождаются в ответ на гормональный сигнал (например, от вазопрессина или тиротропина) под действием специфической мембраносвязанной фосфолипазы С. В результате последовательных реакций образуются два потенциальных вторичных мессенджера – диацилглицерол и инозитол-1,4,5-трифосфат.
Таким образом, представленные данные о вторичных мессенджерах свидетельствуют о том, что каждой из этих систем посредников гормонального эффекта соответствует определенный класс протеинкиназ, хотя нельзя исключить возможности существования тесной связи между этими системами. В результате меняется не только активность, но и регуляторные и каталитические свойства многих ферментных систем клетки: ионных каналов, внутриклеточных структурных элементов и генетического аппарата.
2)Реализация эффекта после проникновения гормона внутрь клетки
В этом случае рецепторы для гормона находятся в цитоплазме клетки. Гормоны этого механизма действия в силу своей липофильности легко проникают через мембрану внутрь клетки-мишени и связываются в ее цитоплазме специфическими белками-рецепторами. Гормон-рецепторный комплекс входит в клеточное ядро. В ядре комплекс распадается, и гормон взаимодействует с определенными участками ядерной ДНК, следствием чего является образование особой матричной РНК. Матричная РНК выходит из ядра и способствует синтезу на рибосомах белка или белка-фермента. Так действуют стероидные гормоны и производные тирозина — гормоны щитовидной железы. Для их действия характерна глубокая и длительная перестройка клеточного метаболизма.
Известно, что эффект стероидных гормонов реализуется через генетический аппарат путем изменения экспрессии генов. Гормон после доставки с белками крови в клетку проникает (путем диффузии) через плазматическую мембрану и далее через ядерную мембрану и связывается с внутриядерным рецептором–белком. Комплекс стероид–белок затем связывается с регуляторной областью ДНК, с так называемыми гормончувствительными элементами, способствуя транскрипции соответствующих структурных генов, индукции синтеза белка de novo и изменению метаболизма клетки в ответ на гормональный сигнал.
Следует подчеркнуть, что главной и отличительной особенностью молекулярных механизмов действия двух основных классов гормонов является то, что действие пептидных гормонов реализуется в основном путем посттрансляционных (постсинтетических) модификаций белков в клетках, в то время как стероидные гормоны (а также тиреоидные гормоны, ретиноиды, витамин D3-гормоны) выступают в качестве регуляторов экспрессии генов.
Инактивация гормонов происходит в эффекторных органах, в основном в печени, где гормоны претерпевают различные химические изменения путем связывания с глюкуроновой или серной кислотой либо в результате воздействия ферментов. Частично гормоны выделяются с мочой в неизмененном виде. Действие некоторых гормонов может блокироваться благодаря секреции гормонов, обладающих антагонистическим эффектом.
– Конец работы –
Эта тема принадлежит разделу:
Гены и ДНК
На сайте allrefs.net читайте: "Гены и ДНК"
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Механизм влияния гормонов на организм
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?
Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.018 сек.
Новости и инфо для студентов